There has, in the last few years, been an increase in the amount of research
de voted to the problem of attention, which has been summarized in Broadbent's
(1958) important work. Whilst psychologists have been investigating the behavioral
aspects of attention, suggestive evidence has also been found by neurophysiologists.
We feel that it would be useful at this time to consider the theoretical implications
of some of this research.
Our paper is divided into three parts. In the first we consider some of the behavioral findings on attention. In the second a system is proposed to account for various features of this behavior. Although we do not consider it necessary to identify a system of this type with particular neural structures (see Deutsch, 1960) since a machine embodying such a system would also display the behavior we wish to explain, we do, however, venture some tentative hypotheses concerning the neural identification of the proposed system.
By: J. Anthony Deutsch and D. Deutsch -- From J. Anthony Deutsch and D. Deutsch, "Attention: Some Theoretical Considerations," Psychological Review, 1%3, 70, 80-90. Reproduced with permission of the authors and publisher.
However alert or responsive we may be, there is a limit to the number of things to which we can attend at any one time. We cannot, for instance, listen effectively to the conversation of a friend on the telephone if someone else in the room is simultaneously giving us complex instructions as to what to say to him. And this difficulty in processing information from two different sources at the same time occurs even if no overt response is required. This phenomenon of selective attention has been investigated in a number of experiments. The most important of these deals with the processing of information emitted simultaneously by two separate sound sources (Broadbent, 1954; Cherry, 1953; Spieth, Curtis, and Webster, 1954). Two problems arise from the results of such experiments. The first is how different streams of information are kept distinct by the nervous system, and how a resultant babel is thereby avoided. The second is why only one of the messages (once it has been kept distinct and separate) is dealt with at any one time. A proposed solution to the first problem, based on experiments in which two messages were fed simultaneously one to each ear, was that the messages were kept distinct by proceeding down separate channels (such as different neural path ways). Nor was it difficult for Broadbent (1958) to extend such a notion to other cases. It had been shown in numerous experiments that we are enabled to listen to one of two simultaneous speech sequences while ignoring the other, by selecting items for attention which have some feature or features in common, such as their frequency spectra (Egan, Carterette, and Thwing, 1954; Spieth et al., 1954) and their spatial localization (Hirsch, 1950; Poulton, 1953; Webster and Thompson, 1954). It was supposed that relatively simple mechanisms were responsible for segregation according to these categories, though the principles of their operation were not made clear.
Broadbent's (1958) answer to the second problem, of how one message is admitted to the exclusion of others, followed from the notions we have already considered. It was proposed that there was a filter which would select a message on the basis of characteristics toward which it had been biased and allow this message alone to proceed to the central analyzing mechanisms. In this way, messages with other characteristics would be excluded and so the total amount of discrimination which would have to be performed by the nervous system would be greatly decreased. Whole complex messages could be rejected on the sole basis of possessing some simple quality, and no further analysis of them would occur.
However, it seems that selection of wanted from unwanted speech can be performed on the basis of highly complex characteristics. For instance, Peters (1954) found that if an unwanted message is similar in content to the wanted ouv, it produces more interference with the adequate reception of the latter than if it is dissimilar to it. This shows that the content of the two messages is analyzed prior to the acceptance of one and rejection of the other. Gray and Wedderburn (1960) have also found that when speech was delivered to subjects in both ears simultaneously, such that a meaningful sequence could be formed by choosing syllables or words alternately from each ear, the subjects reported back the meaningful sequence rather than the series of words or syllables presented to one ear or the other. Treisman (1960) presented two messages, one to each ear, and subjects were asked to repeat what they heard on one ear. The messages were switched from one ear to the other in the middle and it was found that subjects tended to repeat words from the wrong ear just after the switch. "The higher the transition probabilities in the passage the more likely they were to do this" (Treisman, 1960). Other evidence, indicating that complex discriminations would be required of the filter, has been produced by experiments concerning the selection of novel stimuli, for which function Broadbent (1958) assumes the filter to be responsible. Sharpess and Jasper (1956), studying habituation to auditory stimuli in cats, found that habituation, both behavioral and EEG, was specific not only to the frequency of sound presented, but also to the pattern in which a combination of frequencies was presented. Evidence for human subjects is presented by Sokolov (1960) and Voronin and Sokolov (1960), who report that when habituation has been established to a group of words similar in meaning but different in sound, then arousal occurred to words with a different meaning. Behavioral data on the arousal of curiosity in rats upon the presentation of novel visual patterns are reported by Thompson and Solomon (1954). Such evidence as the above would require us, on filter theory, to postulate an additional discriminative system below or at the level of the filter, perhaps as complex as that of the central mechanism, to which information was assumed to be filtered.
Howarth and Ellis (1961) have presented an ingenious experimental argument to show that the same discriminatory mechanism functions in normal perception and when, on filter theory, the discrimination would have to be performed at the level of the filter. The case they put forward is as follows. Moray (1959) had shown that if a subject is listening selectively to one channel and ignoring the other, calling his name on the rejected channel will on a certain pro portion of instances cause him to switch his attention to this channel. This was explained by assuming that the subject's name had a higher priority for the filter than the message to which he had been attending. Oswald, Taylor, and Treisman (1960) in a well-controlled experiment reported that during sleep a subject tends to respond selectively to his own name. Howarth and Ellis (1961) went on to show that the subject's name has a significantly lower threshold than other names when the subject is required to listen normally and there is masking by noise. After analyzing quantitatively their results and those obtained by Oswald et al. and Moray, they (Howarth and Ellis, 1961) conclude that, There is, therefore, a very impressive amount of agreement among these three very different experiments concerning the relative intelligibility of one's own name. It seems an obvious conclusion to suppose that the same pattern-analyzing mechanism is required to account for behavior during dichotic listening or during sleep . . . as during ordinary listening under noise. Thus although Broadbent's (1958) filter provides an ingenious explanation of the selection of messages by means of simple and few discriminations, such as which ear is being stimulated, it becomes less attractive as an explanation of those cases where complex and many discriminations, discussed above, are needed.
If we may identify levels in filter theory with neural levels, then there is also evidence against a two-level system to account for novelty and habituation on neurological grounds. Sharpless and Jasper (1956) found that specificity of habituation to tonal pattern was destroyed by bilateral regions of cortex concerned with audition. It is known from other work (Goldberg, Diamond, and Neff, 1958) that sound pattern discrimination is a cortical function. On the other hand, frequency specific habituation was maintained with Sharpless and Jasper's lesions and it has been shown that frequency discrimination can be taught to animals without these cortical areas (Goldberg et al., 1958). This shows, first, that the level at which habituation occurs is not the same for both pattern and tone, and second, that the destruction of the level which is essential to normal functioning also destroys an animal's ability to habituate. This renders it plausible to assume that the mechanism responsible for habituation is not on a different level from that responsible for other learning and discrimination.
THEORETICAL CONSIDERATIONS
This review of the behavioral evidence leads us to the probable conclusion that a message will reach the same perceptual and discriminatory mechanisms whether attention is paid to it or not; and such information is then grouped or segregated by these mechanisms. How such grouping or segregation takes place is a problem for perceptual theory and will not concern us here. We may suppose that each central structure which is excited by the presentation of a specific quality or attribute to the senses, is given a preset weighting of importance. The central structure or classifying mechanism with the highest weighting will transfer this weighting to the other classifying mechanisms with which it has been grouped or segregated.
The main point with which we are concerned is the following. Given that there is activity in a number of structures, each with a preset weighting of importance, how might that group of structures with the greatest weighting be selected? Or, in behavioral terms, how might the most important of a group of signals be selected? Any system which performs such a function must compare all the incoming signals in importance. This could be done by comparing each in coming signal continuously to every other incoming signal and deciding which is the most important by seeing which signal has no other signal which exceeds it in the physical dimension by which "importance" is represented. But a small amount of reflection will suffice to show that such a system is very uneconomical.
Each possible incoming signal must have a provision in the shape of numerous comparing mechanisms, through each of which it will be connected to all other possible signals. So that as the number of possible signals increases, the number of mechanisms to compare them all against each other will increase at an enormous rate. If the same comparing mechanisms are to be shared by pairs of signals then the time to reach a decision will increase out of all bounds.
However, there is a simpler and more economical way to decide that one out of a group of entities is the largest. Suppose we collect a group of boys and we wish to decide which is the tallest. We can measure them individually against each other and then select the boy in whom this comparison procedure never yielded the answer "smaller." This is like the system outlined above. The decision smaller will be made in this case when we lower a horizontal plane or ruler down on the heads of two boys. The boy whose head is touched by this instrument is declared to be larger and the other boy smaller. But such a procedure is cumbersome because there are many pairs of boys and we must scan through many records of individual boys before we can select the tallest. We could, of course, argue that a simpler solution would be to use an absolute measure of height, such as a ruler with feet and inches inscribed on it. But this procedure is not really simpler. Each boy must be compared against the ruler, and then the measurements themselves must be compared against each other in much the same way as the boys were to decide on the larger and smaller in each couple.
If we are simply interested in finding the tallest boy, then an alternative procedure may be used. Suppose we collect our group below our board which is horizontal and travels lightly up and down, and then ask all our group to stand up below it. Then the boy whose head touches the board when the whole group is standing up will be the tallest boy in the group. If then we call him out, the board will sink until it meets the head of the next tallest individual. If we intro duce some other boys into the group, then if there is a taller boy in this group the board will be raised until it corresponds to his height. In such a system only the tallest individual will make contact with the board, and so he will himself have an immediate signal that he is the tallest boy.
Now suppose that instead of boys, we have signals, not varying in height, but in some other dimension (which we may continue to call "height") which corresponds to their importance to the organism. Suppose that each signal as it arrives is capable of pushing some "level" up to its own "height" (the height determined by its importance), then the most important signal arriving at any particular time will determine this level, analogous to the horizontal board in our example. It will then be the case that any signals which arrive then or after and are of lesser importance and so of smaller height will be below this level. however, if the signal of greater height ceases to be present, then the level will sink to the height reflecting the importance of one of the other signals which is arriving.
If we suppose that only signals whose height corresponds to the height of the level switch in further processes, such as motor output, memory storage, and whatever else it may be that leads to awareness, we have the outline of a system which will display the type of behavior we associate with attention. Only the most important signals coming in will be acted on or remembered. On the other hand, more important signals than those present at an immediately preceding time will be able to break in, for these will raise the height of the level and so displace the previously most important signals as the highest.
So far we have omitted any discussion of the role of general arousal in selective attention. Without such arousal, usually (but not invariably, Bradley and Elkes, 1953; Gestaut, 1954) indicated by characteristic patterns on the electro encephalogram, awareness of and behavioral responsiveness to peripheral stimulation are absent. Some degree of general arousal is thus necessary for attention to operate. Furthermore, individuals when aroused will attend to any incoming message, provided that it is not concomitant with a more important one, whereas when asleep they will only respond to very "important" messages, such as a per son's own name (Oswald et al., 1960) or, in the case of a mother, the sound of her infant crying. And when drowsy, though responsive to a larger range of stimuli than when asleep, subjects will tend to "miss" signals which they would notice when fully awake.
The system which takes this into consideration is schematically represented in the diagram (Figure 1). Any given message will only be heeded if the horizontal line (Y) representing the degree of general arousal meets or crosses the vertical line, the height of which represents the "importance" of the message.
Whether or not alerting will take place then depends both on the level of general arousal and on the importance of the message. Attention will not be paid to Message b though it is the most important of all incoming signals, when the level of general arousal is low (Position X). When the level of general arousal is at Z, which is very high, attention could be paid to all signals a, b, c, d, and e. In fact, attention is paid only to b as a result of the operation of the specific alerting mechanism.
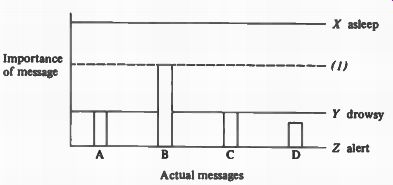
FIGURE 1. Diagram to illustrate operation of proposed system. (The interrupted
horizontal line (1) represents the "level" of importance in the
specific alerting system which is raised and lowered according to the incoming
messages. The solid horizontal lines represent levels of general arousal.
At X, the organism is asleep, and none of the actual messages produce alerting.
At Y, the organism is drowsy, and only some incoming messages produce alerting.
At X, the organism is awake. All messages could he alerted to, but the specific
alerting system allows only h to be heeded.)
Further, it is supposed that a message will increase the level of general arousal in proportion to its importance and for various lengths of time in proportion to its importance, so that messages which would not have been heeded be fore will command attention if they follow in the wake of a more important message.
The mechanism whereby the weighting of importance of messages is carried out is given by Deutsch's (1953, 1956, 1960) theory of learning and motivation, and will be only briefly summarized here, since it is not the main point of the paper. It is assumed that on exposure to a succession of stimuli, link-analyzer units responsive to these stimuli will be connected together. Certain primary links, when stimulated by physiological factors, generate excitation, and this is passed on from link to link along the connections established by experience. Each link-analyzer unit will receive excitation depending first, on the state of the primary links to which it is connected, either directly or indirectly, and second, on the "resistance" of such a connection, which is determined by past learning. It is assumed that the amount of such excitation arriving at a link-analyzer unit deter mines both its threshold of excitability by incoming stimuli (leading to an increased readiness to perceive a stimulus whether it is there or not) and the ranking of importance of such a stimulus (e.g., Lawrence, 1949, 1950). We should predict from this theory an inverse correlation between the attention-getting or distracting value of a stimulus when attention is being paid to another, and its threshold (regarded as the likelihood of its being reported by a subject when he is asked to say what he perceives). We should also expect that stimuli which have a high importance weighting should more often be mistakenly perceived when similar stimuli are present.
NEUROPHYSIOLOGICAL CORRELATES
We may ask how the suggested system would fit what is known of the physiological substrate of attentive behavior. One of the salient features of the system as proposed is that it assumes that all sensory messages which impinge upon the organism are perceptually analyzed at the highest level. It would therefore be of relevance to discuss the group of neuro-physiological experiments, the results of which have been claimed to demonstrate a neural blockage of "rejected" messages at the lower levels of the primary sensory pathways. Hernández-Péon, Scherrer, and Jouvet (1956) showed that the evoked response at the dorsal cochlear nucleus to clicks was reduced by the presentation of "distracting" olfactory and visual stimuli. A similar effect was found in the visual pathways (Hernán dez-Péon, Guzman-Flores, Alcarez and Fernandez-Guardiola, 1957). Stimulation of the reticular formation could produce similar results, and it was supposed that such stimulation was treated as the presentation of a distracting stimulus. It has also been demonstrated by various workers (e.g., Galambos, Sheatz, and Vernier, 1956; Hernández-Péon and Scherrer, 1955) that responses to auditory clicks re corded from the dorsal cochlear nucleus (as well as other placements) diminish with repetition. Habituation to photic stimuli has been demonstrated for the retina ( Palestini, Davidovich, and Hernández-Péon, 1959) and for the olfactory bulb ( Hernández-Péon, Alcocer-Cuaron, Lavin, and Santibafiez, 1957). It was therefore proposed that during inattention to a signal (either by distraction or habituation) information concerning this signal was blocked at the level of the first sensory synapse by means of "afferent neuronal inhibition." Recently, however, evidence has been produced indicating, at least for the visual and auditory pathways, that such changes in the evoked potential were due to peripheral factors, and represented simply a decrease in the effective intensity of the stimulus.
Hugelin, Dumont, and Paillas (1960) report that when the middle ear muscles were cut stimulation of the reticular formation would not cause a diminution in the amplitude of the evoked responses. They report further that such contractions of the middle ear muscles which result from reticular stimulation produce a mean diminution of microphonic potentials of less than 5 decibels. The reduction in sensation brought about by these means therefore appears unimportant. Naquet, Regis, Fischer-Williams, and Fernandez-Guardiola (1960) found that if the size of the pupil were fixed by local application of atropin the evoked potential re corded from placements below the cortex demonstrated a consistent amplitude.
The above findings do not, however, apply to changes in the cortical evoked response during distraction or habituation. Moushegian, Rupert, Marsh, and Galambos (1961) found that in animals in which the middle ear muscles had been cut, cortical evoked responses to clicks still demonstrated diminution during habituation and distraction, and amplification when the clicks were associated with puffs of air to the face. Naquet, in the experiment quoted above, reports that application of atropin to the pupil did not prevent a variation in cortical evoked responses, which diminished during de-synchronization and were enhanced during synchronization of electrical rhythms, also changing in morphology.
Reports of changes in cortical evoked responses during habituation and dis. traction are many and varied, and it would be impossible in this space to de scribe the field in detail. Certainly disagreement exists over what occurs as well as over its interpretation. For instance, Horn (1960), recording flash evoked responses in the visual cortex of cats when resting, and when watching a mouse, found that the responses were reduced in amplitude when the cat was watching the mouse; when it ignored the mouse, responses remained of high amplitude.
Further, after a series of tone-shock combinations, it was noted that the evoked response to flash was reduced after a series of tones only if there was "some visual searching component in the cat's response to the acoustic stimuli." Horn argues that attenuation of evoked responses in the cortex might be correlated with greater sensitivity in the appropriate region, rather than signifying a reduction in incoming information. However, other recent experimenters continue to maintain that evoked responses diminish in amplitude when attention is not being paid to the test stimulus. Garcia-Ausst, Bogacz, and Vanzulli (1961), re corded scalp visual evoked responses in human subjects (who were able to give introspective reports) during presentation of flash stimuli. They report, When the stimulus is significant and therefore attention is paid to it, the response is relatively simple and widespread. When, on the other hand, the stimulus is not significant and no great attention is paid to it, the response is reduced, complex, and localized.
It would seem that changes in the evoked potential at the cortex do indeed take place during habituation and attention shifts; but that what those changes exactly are, and what they represent, is not yet clear.
We should indeed expect, on the above theory of attention, changes in the cortical evoked potential when attention is being paid to a stimulus, reflecting the activation of various processes, such as motor output and memory storage. Pertinent to this assumption is the discovery by Hubel, Henson, Rupert, and Galambos (1959) of what they term "attention" units in the auditory cortex. By the use of microelectrodes implanted in un-anesthetized and unrestrained cats, they obtained records from units which responded only when the animal was "paying attention" to the sound source. These attention units appeared to be both interspersed amongst the others and segregated from them. We may venture to interpret these results by supposing that the units in question formed part of the systems, discussed above, responsible for the appropriate motor response to stimulation or the committing of items to memory, and so forth, or that they lay on the pathway to these systems. Thus they would be inactive even if impulses evoked by auditory stimulation were reaching the cortex, provided that the animal was not also attending to the stimuli.
There is another theoretical assumption for which we might reasonably seek a neuro-physiological counterpart. We suppose that a selection of inputs from a variety of sources takes place by comparison with a fluctuating standard. This implies the existence of an undifferentiated structure with widespread connections with the rest of the central nervous system. We are tempted, on account of the evidence for the diffuseness of its input, to identify the brain stem reticular formation as this particular structure. Potentials may be evoked throughout this structure by excitation of various sensory systems (French, Amerongen, and Magoun, 1952; Starzl, Taylor, and Magoun, 1951), and various cortical structures (Bremer and Terzuolo, 1954; French, Hernández-Péon, and Livingston, 1955). Occlusive and facilitatory interaction between responses evoked in the reticular formation from very different sources have further been observed (Bremer and Terzuolo, 1952, 1954; French et al., 1955). Single unit studies demonstrating a convergence of input from several sources have also been reported (Amassian, 1952; Amassian and De Vito, 1954; Hernández-Péon and Hagbarth, 1955; Scheibel, Scheibel, Mollica, and Moruzzi, 1955). A similar conclusion, that the reticular formation is capable of acting as a nonspecific system, can be based on neuro-anatomical evidence. Scheibel and Scheibel (1958) state on the basis of their extensive histological study:
. . . the degree of overlap of the collateral afferent plexuses is so great that it is difficult to see how any specificity of input can be maintained, rather it seems to integrate and vector a number of inputs.
We have also postulated that the fluctuating level correlates with states of arousal. Again the brain stem reticular formation seems well suited to fulfill this function. Its importance in the regulation of states of arousal has been demonstrated both through work involving lesions (Bremer, 1935; French, 1952; French and Magoun, 1952; Lindsley, Schreiner, Knowles, and Magoun, 1949) and stimulation of this structure (Moruzzi and Magoun, 1949; Segundo, Arana, and French, 1955). Recently Moruzzi (1960) has shown that the lower brain stem may play an important role in the initiation of sleep. It also seems likely that the thalamic reticular system is involved in the regulation of states of arousal. Large bilateral lesions of the anterior portion of this system may produce coma analogous to that produced by lesions of the mid-brain (French et al., 1952) although the depth of coma so produced is less profound. Stimulation of portions of this system has also been shown to produce either sleep or arousal depending on the parameters of stimulation (Akimoto, Yamaguchi, Okabe, Nakagawa, Nakamura, Abe, Torii, and Masahashi, 1956; Hess, 1954). The work of Adametz (1959), Chow and Randell (1960), and Doty, Beck, and Kooi (1959), who demonstrated that with different operational techniques and with assiduous nursing care massive lesions of the mid-brain-reticular formation need not produce coma, should, however, be considered. Chow, Dement, and Mitchell (1959) found also that massive lesions in the thalamic reticular system need not produce coma. Until reasons for these discrepant results are found we must regard our conclusions as to the role of the reticular system in attention as tentative.
Whatever the explanation of the findings on lesions in the reticular formation may turn out to be, it seems that, if we are right, some diffuse and nonspecific system is necessary as a part of the mechanism sub-serving selective attention. Such a system should be found to have afferent connections from all discriminatory and perceptual systems. Through these connections it should be influenced to take up a variety of levels; the level at any one time corresponding with the level of the "highest" afferent message from the discriminatory mechanisms. On its efferent side such a nonspecific system should again be connected with all discriminatory and perceptual mechanisms. Through such connections it would signal to them its own level. If this level of the nonspecific system was above that of a particular discriminatory mechanism, no registration in memory or motor adjustment would take place, if such a discriminatory mechanism was stimulated. Consequently, only that discriminatory mechanism being activated whose level was equal to that of the diffuse system would not be affected. In this way the most important message to the organism will have been selected.
REFERENCES
ADAMETZ, J. H. Rate of recovery of functioning in cats with rostral reticular lesions. I. Neurosurg., 1959, 16, 85-98.
AKIMOTO, H., YAMAGUCHI, M., OKADE, K., NAKAGAWA, T., NAKAMURA, I ., ABE, K., TORII, H., & MASAHASHI, I. On the sleep induced through electrical stimulation of dog thalamus. Folia psychiat. neurol. lap., 1956, 10, 117-146.
ANIASSIAN, V. E. Interaction in the somatovisceral projection system. Proc. Ass. Res. Nerv. Ment. Dis., 1952, 30, 371-402.
AMASSIAN, V. E., & DEVrro, R. V. Unit activity in reticular formation and nearby struc tures. J. Neurophysiol., 1954, 17, 575-603.
BRADLEY, P. B., & axes, J. The effect of atropine, hyoscyamine, physostigmine and neo stigmine on the electrical activity of the brain of the conscious cat. J. Physiol., 1953, 120, 14-15.
BREMER, F. Cerveau isolé et physiologie du sommeil. CR Soc. Biol., Paris, 1935, 118, 1235 1242.
BREMER, F., & Unlit°, C. Rôle de l'écorce cérébrale dans le processus physiologique du réveil. Arch. int. Physiol., 1952, 60, 228-231.
BREMER, F . & TERZUOLO, C. Contribution à l'étude des mécanismes physiologiques du main tein de l'activité vigile du cerveau. Interaction de la formation réticulée et de l'écorce cérébrale dans le processus des réveil. Arch. inter. Physiol., 1954, 62, 157-178.
BROADBENT, D. E. The role of auditory localization in attention and memory span. J. exp. Psychol., 1954, 47, 191-196.
BROADBENT, D. E. Perception and Communication. London: Pergamon, 1958.
CHERRY, E. C. Some experiments on the recognition of speech with one and with two ears. J. Acoust. Soc. Amer., 1953, 25, 975-979.
CHOW, K. L ., DEMENT, W. C., & MITCHELL, S. A., in. Effects of lesions of the rostra] thalamus on brain waves and behavior in cats. EEG clin. Neurophysiol., 1959, 11, 107-120.
CHOW, K. L, & RANDELL, W. Learning and EEG studies of cats with lesions in the reticular formation. Paper read at the first annual meeting of Psychonomics Society, Chicago, 1960.
DEUTSCH, J. A. A new type of behavior theory. Brit. J. Psychol., 1953, 44, 305-317.
DEUTSCH, J . A. A theory of insight, reasoning and latent learning. Brit. J. Psychol., 1956, 47, 115-125.
DEUTSCH, J . A. The structural basis of behavior. Chicago: Univer. Chicago Press, 1960.
DOTY, R. W., BECK, E. C., & Kum, R. A. Effect of brain-stem lesions on conditioned responses in cats. Exp. Neurol., 1959, 1, 360-385.
EGAN, J . P., CARTERETTE, E. C., & THWING, E. J . Some factors affecting multi-channel listening. J. Acoust. Soc. Amer., 1954, 26, 774-782.
FRENCH, J. D. Brain lesions associated with prolonged unconsciousness. AMA Arch. Neurol.
Psychiat., 1952, 68, 727-740. F RENCH, J . D., AMERONGEN, F. K., & MAGOUN, H. W. An activating system in brain stem of monkey. AMA Arch. Neurol. Psychiat., 1952, 68, 577-590.
FRENCH, J. D., HERNA NDEZ-K ON, R., & L IVINGSTON, R. B. Projections from cortex to cephalic brain stem (reticular formation) in monkey. J. Neurophysiol., 1955, 18, 74-95.
F RENCH, J . D., & MAGOUN, H. W. Effects of chronic lesions in central cephalic brain stem of monkeys. AMA Arch. Neurol. Psychiat., 1952, 68, 591-604.
GALAbiRos, R., S HEATZ, G., & VERNIER, V. G. Electrophysiological correlates of a conditioned response in cats. Science, 1956, 123, 376-377.
GARCIA-AUSTT, E., B OGACZ, J ., & VANZULLI, A. Significance of the photic stimulus on the evoked responses in man. In Delafresnaye (Ed.), Brain mechanisms and learning. Oxford: Blackwell, 1961. Pp. 603-626.
GASTALIT, H. The brain stem and cerebral electrogenesis in relation to consciousness. In Delafresnaye (Ed.), Brain mechanisms and consciousness. Springfield, Ill.: Charles C Thomas, 1954. Pp. 249-283.
GOLDBERG, J . M., DIAMOND, I . T., & NEFF, W. D. Auditory discrimination after ablation of temporal and insular cortex in cat. Federat. Proc., 1957, 16, 47-48.
GOLDBERG, J . M., DIAMOND, I . T., & NEFF, W. D. Frequency discrimination after ablation of cortical projection areas of the auditory system. Federat. Proc., 1958, 17, 216-255.
GRAY, J. A., & WEDDERBURN, A. A. I . Grouping strategies with simultaneous stimuli. Quart. J. exp. Psychol., 1960, 12, 180-184.
HERNANDEZ- PÉON, R., ALCOCER-CUARON, C., LAVIN, A., & S ANTIBA R EZ, G. Regulación cen trifuga de la actividad électrica del bulbo olfactorio. Punta del Este, Uruguay: Primera Reunión Cientifica de Ciencias Fisiólogicas, 1957. Pp. 192-193.
HERANDEZ- N ON, R., GUZMAN-F LORES, C., ALCAREZ, H., St FERNANDEZ-GUARDIOLA, A. Sen sory transmission in visual pathway during "attention" in un-anaesthetized cats. Acta neurol. Lat.-Amer., 1957, 3, 1-8.
HERNANDEZ- PÉON, R., & HAEBARTH, K. Interaction between afferent and cortically induced reticular responses. J. Neurophysiol., 1955, 18, 44-55.
HERNANDEZ- PON, R., & SCHERRER, H. Habituation to acoustic stimuli in cochlear nucleus. Federat. Proc., 1955, 14, 71.
HERNÁNDEZ- PÉON, R., S CHERRER, H., & Jouyey, M. Modification of electric activity in cochlear nucleus during "attention" in unanaesthetized cats. Science, 1956, 123, 331 332.
Hess, W. R. The diencephalic sleep centre. In J . F. Delafresnaye (Ed.), Brain mechanisms and consciousness. Springfield, Ill., Charles C Thomas, 1954. Pp. 117-136.
HIRSH, I . J. The relation between localization and intelligibility. J. Acoust. Soc. Amer., 1950, 22, 196-200.
HORN, G. Electrical activity of the cerebral cortex of the un-anaesthetized cat during attentive behavior. Brain, 1960, 83, 57-76.
HOWARTH, C. I., & Ems, R. The relative intelligibility threshold for one's own name compared with other names. Quart. J. exp. Psychol., 1961, 13, 236-239.
Ilium., D. H., HENSON, C. O., RUPERT, A., & GALAMBOS, R. Attention units in the auditory cortex. Science, 1959, 129, 1279-1280.
HUGELIN, A., DUMONT, S., & PAILLAS, N. Tympanic muscles and control of auditory input during arousal. Science, 1960, 131, 1371-1372.
LAWRENCE, D. H. Acquired distinctiveness of cues: I. Transfer between discriminations on the basis of familiarity with the stimulus. I. exp. Psychol., 1949, 39, 770-784.
LAWRENCE, D. H. Acquired distinctiveness of cues: H. Selective association in a constant stimulus situation. J. exp. Psychol., 1950, 40, 175-188.
LimnsLev, D. B., SCHREINER, L. H., KNOWLES, W. B., & MAcourt, H. W. Behavioral and EEG changes following chronic brain stem lesions in the cat. EEG clin. Neurophysiol., 1949, 1, 455-473.
MORAY, N. Attention in dichotic listening: Affective cues and the influence of instructions. Quart. J. exp. Psychol., 1959, //, 56-60.
MORUZZI, G. Synchronizing influences of the brain stem and the inhibitory mechanisms underlying the production of sleep by sensory stimulation. Int. J. EEG clin. Neuro physiol., 1960, Suppl. No. 13, 231-256.
MORUZZI, G., & MAGOUN, H. W. Brain stem reticular formation and activation of the EEG. EEG clin. Neurophysiol., 1949, 1, 455-473.
MOUSHEGIAN, G., RUPERT, A., MARSH, J. T., & GALAMBOS, R. Evoked cortical potentials in absence of middle ear muscles. Science, 1961, 133, 582-583.
NAQUET, R., REGIS, H., FISCHER-WILLIAMS, M., & FERNANDEZ-GUARDIOLA, A. Variation in the responses evoked by light along the specific pathways. Brain, 1960, 83, 52-56.
OSWALD, I., TAYLOR, A., & TREISMAN, M. Discrimination responses to stimulation during human sleep. Brain, 1960, 83, 440-453.
PALESTINI, M., DAVIDOVICH, A., & HERNÁNDEZ-PON, R. Functional significance of centrifugal influences upon the retina. Acta neurol. Lat.-Amer., 1959, 5, 113-131.
PETERS, R. W. Competing messages: The effect of interfering messages upon the reception of primary messages. USN Sch. Aviat. Med. res. Rep., 1954, Project No. NM 001 064.01.27.
POULTON, E. C. Two-channel listening. J. exp. Psychol., 1953, 46, 91-96.
SCHEIBEL, M. E., & SCHEIBEL, A. Structural substrates for integrative patterns in the brain stem reticular core. In H. H. Jasper, L. D. Proctor, R. S. Knighton, S. Roberts, W. C. Noshay, C. William, and R. T. Costello (Eds.), Reticular formation of the brain. Boston: Little, Brown, 1958. Pp. 31-55.
SCHEIBEL, M. E., SCHEIBEL, A., MOLLICA, A., & MORUZZI, G. Convergence and interaction of afferent impulses on single units of reticular formation. I. Neurophysiol., 1955, 18, 309-331.
SEGUNDO, J. P., ARANA, R., & FRENCH, J. D. Behavioral arousal by stimulation of the brain in monkey. J. Neurosurg., 1955, 12, 601-613.
SHARPLESS, S., & JASPER, H. Habituation of the arousal reaction. Brain, 1956, 79, 655-678.
SOKOLOV, E. N. Neuronal models and the orienting reflex. In Mary A. B. Brazier (Ed.), The central nervous system and behavior: Transactions of the third conference. New York: Josiah Macy, Jr. Foundation, 1960, Pp. 187-276.
SPIETH, W., CURTIS, J. F., & WEBSTER, J. C. Responding to one of two simultaneous messages. J. Acoust. Soc. Amer., 1954, 26, 391-396.
STARZL, T. E., TAYLOR, C. W., & MACOUN, 11 W. Collateral afferent excitation of reticular formation of brain stem. J. Neurophysiol., 1951, 14, 479-496.
THOMPSON, W. R., & S OLOMON, L . M. Spontaneous pattern discrimination in the rat. J. comp. physiol. Psychol., 1954, 46, 281-287.
TREISMAN, A. M. Contextual cues in selective listening. Quart. J. exp. Psychol., 1960, 12, 242-248.
VORONIN, L. G., & Sox()Lov, E. N. Cortical mechanisms of the orienting reflex and its relation to the conditioned reflex. Int. J. EEG clin. Neurophysiol., 1960, Suppl. No. 13, 335-346.
WEBSTF.R, J. C., & THOMPSON, P. 0. Responding to both of two overlapping messages. J. Acoust. Soc. Amer., 1954, 26, 396-402.
Also in Part 3:
- Communication Theory: Decoding-Encoding
- Perception and Communication: A Transactional View
- Perception, Attention, and Consciousness
- Attention: Some Theoretical Considerations
- The Human Receiving System